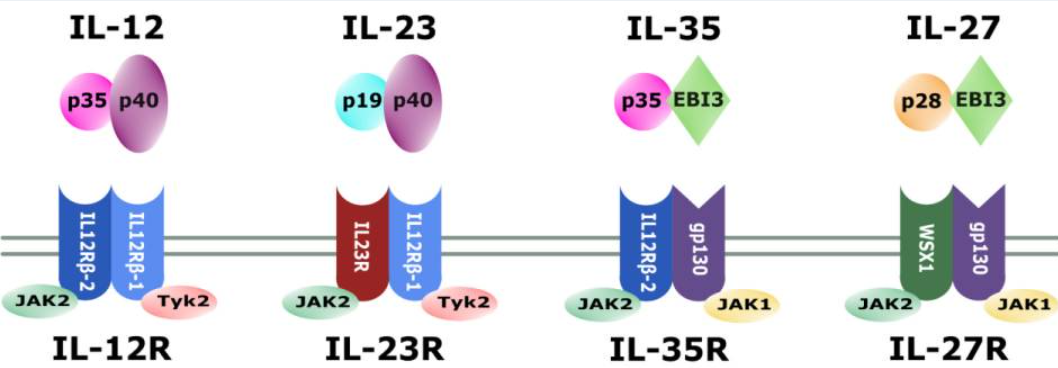
IL-12的分子结构
IL-12是一种异源二聚体细胞因子,由α链p35(约35 kDa)和β链p40(约40 kDa)组成。这两个亚基分别由不同基因编码(人类基因位于第3号和5号染色体)。值得注意的是,p35本身不具备独立分泌的能力,必须与p40结合后才能正确折叠并分泌为生物活性的IL-12异质二聚体(通常称为IL-12p70)。IL-12家族其它成员也采用类似结构:例如IL-12p40与另一α链IL-23p19可组成IL-23,而p35也可与EBI3(EB病毒诱导基因3)结合形成免疫抑制性细胞因子IL-35。这种共享亚基机制使得IL-12与IL-23等细胞因子在结构上具有亲缘关系。
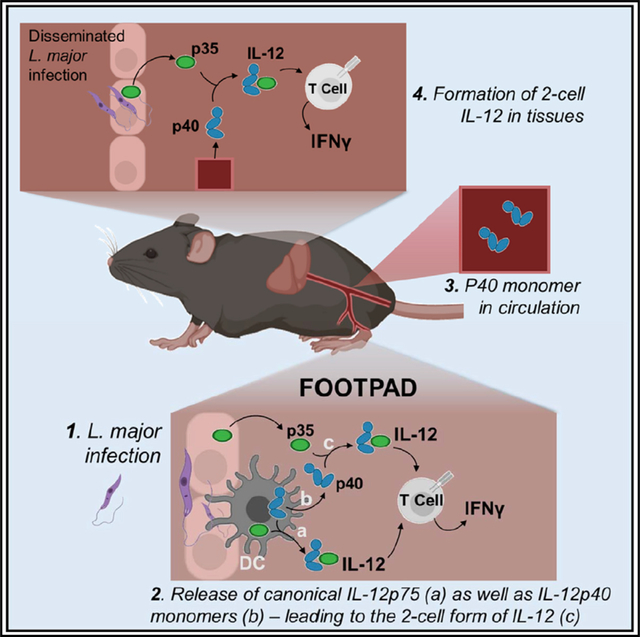
IL-12p40亚基在血清和细胞培养上清中常以过量单体形式出现,并能形成p40-p40同二聚体。研究表明,p40同二聚体可拮抗IL-12p70促进IFN-γ产生的活性,尽管其生理意义尚未完全明了。p40单体过量分泌可能参与调节免疫平衡,而p35的表达则更加广泛,但仅在与p40结合后才能发挥作用。
IL-12受体的组成与表达分布
IL-12的细胞表面受体由两个跨膜亚基构成:IL-12Rβ1和IL-12Rβ2。其中IL-12Rβ1与IL-23受体共用(IL-23信号复合体由IL-12Rβ1和专属的IL-23R构成),而IL-12Rβ2则特异性地与IL-12结合形成高亲和力受体。IL-12Rβ1与IL-12Rβ2同属于gp130家族,其胞外结构域与gp130蛋白具有同源性。在免疫细胞中,两种亚基主要表达在活化的T淋巴细胞和NK细胞表面。活化状态下的B细胞也可表达IL-12受体:在IFN-γ和IL-12共同作用下,人B细胞上调IL-12Rβ2表达,并在IL-12刺激下发生STAT4磷酸化,表明B细胞在特定条件下也能响应IL-12信号。值得一提的是,IL-12Rβ1的亲和力较低(Kd ~2–6 nM),每细胞表达数量较多(约1000–5000个受体),而IL-12Rβ2则为高亲和力受体(Kd ~5–20 pM),表达数量相对较少(约100–1000个)。
在IL-12信号传导中,IL-12Rβ1和IL-12Rβ2的胞内区分别结合不同的Janus激酶:IL-12Rβ1主要结合Tyk2,而IL-12Rβ2结合JAK2。IL-12与受体结合后,受体相关的JAK2和TYK2被活化并发生自磷酸化,进而磷酸化受体胞内区上的酪氨酸。IL-12Rβ2上关键的磷酸化位点(如Y800)为STAT4提供结合位点,形成信号平台。此外,STAT4可被激酶家族中的p38 MAPK磷酸化(丝氨酸位点),这一修饰对STAT4的转录活性和IFN-γ产生至关重要。此外,IL-12还可激活PI3K/Akt通路,促进T细胞增殖。
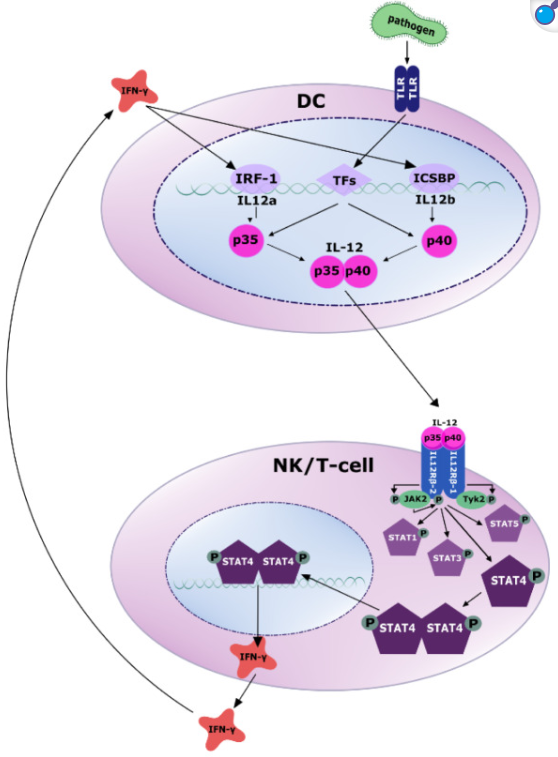
抗原呈递细胞 (APC),例如树突状细胞,通过 Toll 样受体(TLR) 感知 PAMP(病原体相关分子模式)。随后,多种转录因子被激活,诱导 IL-12p35 和 IL-12p40 的转录(更多详情请参阅原文)。分泌的 IL-12 异二聚体与 NK 细胞和 T 细胞上的受体结合,募集酪氨酸激酶 JAK2 和 TYK2,并通过酪氨酸磷酸化激活 JAK2。活化的 JAK2 磷酸化 IL12Rβ2 亚基,进而通过磷酸化激活 STAT4。随后,磷酸化的 STAT4 发生同二聚化或异二聚化,转位至细胞核,并通过结合靶 DNA 调控基因转录。主要靶基因是 IFN-γ,其反过来通过 IRF-1 和 ICSBP 诱导 IL-12 产生的转录激活。
IL-12信号的负反馈调控
为防止过度的炎症反应,IL-12信号通路受到多种负反馈机制的制约。抑制性细胞因子信号分子SOCS-3是IL-12负调节的关键分子之一。SOCS-3的SH2结构域能够识别并结合磷酸化的IL-12Rβ2上的Y800位点,从而竞争性阻断STAT4与受体的结合。研究显示,在高表达SOCS-3的Th2细胞中,IL-12介导的STAT4活化被强烈抑制,表明SOCS-3在Th2细胞中防止IL-12信号介导的Th1极化。除SOCS-3外,其他SOCS蛋白或蛋白酪氨酸磷酸酶也可能参与IL-12信号的调控,但SOCS-3对IL-12Rβ2的抑制作用是已知的经典机制。此外,IL-12能诱导的IFN-γ和其它下游效应也可间接反馈调节IL-12的生成,但主要的细胞内负反馈仍归功于SOCS-3等分子的作用。
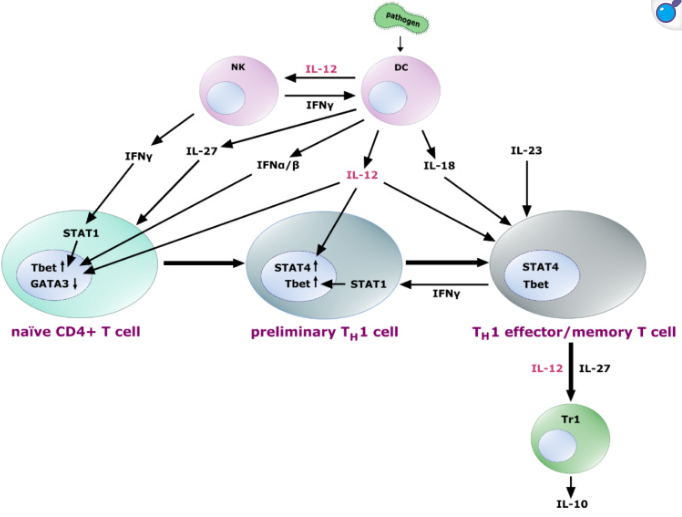
暴露于 IL-27 的幼稚 T 细胞表达 IL-12 受体异二聚体,使细胞对 IL-12 的影响敏感。IL-12 与来自 NK 细胞和反馈回路的 IFN-γ(IFN-α和 IFN-β)共同诱导转录因子 Tbet的上调和 GATA3的下调,从而初步实现 TH1 分化。进一步暴露于 IL-12 会导致这些早期 TH1 细胞中 STAT4 的上调,随后它们分化为 TH1 效应 T 细胞和记忆 T 细胞。IL-18和 IL-23 有助于 TH1 细胞效应功能的固定、扩增和维持。分化的 TH1 细胞产生 IFN-γ,通过 STAT1 增强 Tbet 的表达,从而形成正反馈回路。此外,它们能够促进 IL-10 分泌 Tr1 细胞,这一过程可能由 IL-12 和 IL-27 信号介导。
IL-12的上游诱导因素
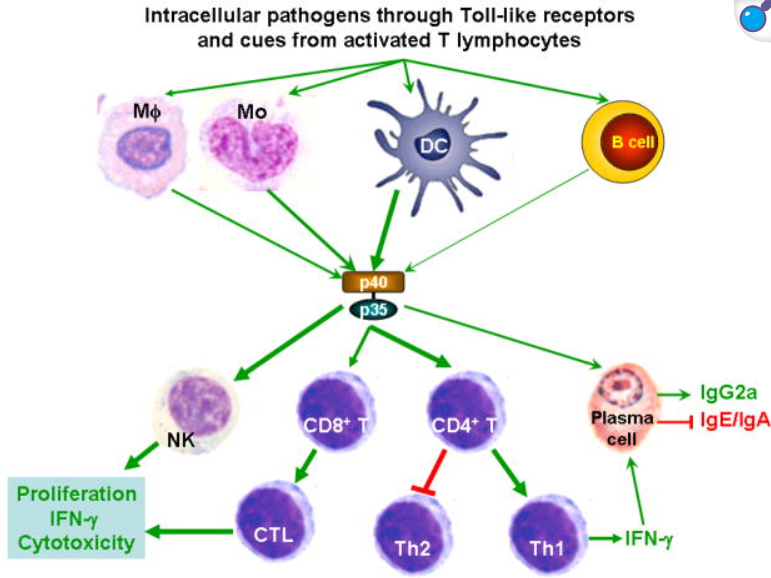
IL-12的产生主要由树突状细胞和巨噬细胞介导,这些抗原呈递细胞在受到病原体相关分子模式(PAMPs)刺激后通过TLR通路激活相关转录因子来合成IL-12。经典的例子包括细胞毒性链霉菌毒素(CpG)激活TLR9、脂多糖(LPS)激活TLR4或其他TLR激活均能诱导IL-12的表达。分子机制上,TLR3/4活化时细胞内IRF-3可结合IL-12p35基因启动子中的ISRE元件,促进p35转录。同时,NF-κB和AP-1等转录因子也参与IL-12p40亚基的启动子激活(相关文献显示IRF-5等因子可诱导p40的表达,未于此重点讨论)。在炎症环境中,IFN-γ是IL-12产生的强烈增强因子:IFN-γ能诱导转录因子IRF-1和ICSBP(IRF-8),分别作用于IL-12p35和p40基因,形成正反馈环路。结果是IL-12激发的初始IFN-γ分泌又进一步提高后续IL-12的生成。此外,炎性细胞因子IL-18虽然自身对IFN-γ诱导作用有限,但与IL-12协同时可显著放大IFN-γ的产生:IL-18通过STAT4和AP-1增强IFN-γ基因转录,特别在巨噬细胞或B细胞等非经典IFN-γ来源细胞中尤为重要。总之,IL-12的上游诱导涉及TLR-介导的NF-κB/IRF信号通路,以及IFN-γ/IL-18等炎症因子的正反馈放大效应,确保IL-12在免疫应答中的及时释放。
IL-12的下游生物学效应
IL-12的主要生物学作用是激活先天和适应性免疫效应细胞,从而引导免疫应答向Th1型极化。IL-12可直接刺激自然杀伤(NK)细胞和T淋巴细胞,使其大量分泌IFN-γ并增强细胞毒性。在CD4+ T细胞中,IL-12通过STAT4信号促进幼稚T细胞分化为Th1细胞,使Th1分泌表型(如高水平IFN-γ、T-bet表达)得到建立。IFN-γ反过来上调MHC分子、促进巨噬细胞杀菌效能,并促进IL-12Rβ2的表达,形成正反馈,进一步巩固Th1极化。IL-12还能抑制Th2细胞相关因子的表达,使免疫应答更加偏向细胞介导的抗原特异性免疫。在CD8+ T细胞中,IL-12作为“信号3”参与初始激活,可促进效应CD8+ T细胞的增殖和细胞毒作用,并对其向长期记忆细胞转化具有关键作用。研究表明,在抗原初次遇到早期,如果CD8+ T细胞获得IL-12或Ⅰ型干扰素信号,其后可形成具有保护性效应的记忆T细胞群体。此外,IL-12也能间接促进B细胞的辅助活性,如通过诱导Th1细胞表位帮助B细胞产生IgG2a(未于此详细展开)。总体而言,IL-12通过增强NK细胞和CTL功能、驱动Th1细胞分化并促进记忆T细胞生成,实现先天免疫与适应性免疫的桥接。
不同微环境中的IL-12调控与细胞选择性效应
IL-12信号的效果高度依赖于组织和炎症微环境的特点。在感染性炎症微环境中(如细菌、寄生虫感染),IL-12产生通常被显著诱导:巨噬细胞或树突状细胞通过TLR识别病原时大量分泌IL-12,以促进强烈的Th1反应和IFN-γ释放。临床上已知IL-12Rβ1缺陷会严重损害机体对结核分枝杆菌和沙门氏菌等胞内病原的清除能力,突显IL-12在抗感染中的关键作用。相反,在慢性炎症或肿瘤微环境中,IL-12信号经常受到抑制。许多肿瘤细胞通过激活STAT3信号通路和分泌IL-10、TGF-β等抑制性因子来抑制IL-12的生成,同时增强IL-23的产生,使免疫偏向促炎的Th17/IL-17路径。这种IL-12/IL-23平衡的转变利于肿瘤或慢性炎症的持续发展。针对肿瘤免疫治疗研究发现,高剂量IL-12治疗往往伴随机体产生大量IL-10反调节,但IL-12仍能极大增强肿瘤特异性CTL应答。在小鼠肿瘤模型中,IL-12给药可显著延缓肿瘤生长,并诱导肿瘤内部产生抑制血管生成的趋化因子(如IP-10、MIG)。临床试验也观察到IL-12治疗后肿瘤患者外周血和转移灶中肿瘤特异性CD8+ CTL及记忆T细胞显著增多。此外,IL-12的细胞选择性效应也十分明确:例如Th2细胞高水平表达SOCS-3,可阻断IL-12介导的STAT4信号,导致Th2对IL-12响应性很低。相反,Th1细胞则能有效响应IL-12刺激。综上所述,IL-12信号在感染环境中被增强以促进免疫防御,而在肿瘤或慢性炎症中常被负反馈抑制;不同免疫细胞对IL-12反应的差异则决定了其细胞选择性效应。
IL-12在免疫治疗中的应用潜力
鉴于IL-12在连接先天与适应性免疫中的中心作用,它成为免疫治疗的重要靶点之一。在抗肿瘤免疫方面,IL-12能通过增强CTL和NK细胞介导的杀伤以及抑制肿瘤血管生成来发挥作用。研究发现IL-12刺激的IFN-γ可上调肿瘤细胞表面MHC和ICAM-1分子,并诱导内皮细胞产生IP-10/MIG等抗血管生成因子,从而抑制肿瘤生长。小鼠实验显示,IL-12单独或作为疫苗佐剂可显著延长肿瘤模型动物生存期并提高肿瘤特异性免疫应答。例如,将IL-12基因导入肿瘤疫苗中,与常规疫苗联合使用时比单独使用任何一种效果更佳。然而,系统给药IL-12常导致显著炎症毒性,现代策略正探索局部释放(如肿瘤内注射)、基因治疗或与其它检查点抑制剂联合的方案,以优化疗效并减少副作用。此外,IL-12通路的调控也为治疗自身免疫疾病提供了思路,针对IL-12/IL-23p40亚基的抗体(如乌司奴单抗)已用于银屑病和克罗恩病等炎性疾病,说明IL-12信号作为调节节点在免疫治疗中具有双重意义(既可增强抗原反应亦可通过阻断减轻自身炎症)。总体来看,最新研究继续揭示IL-12信号在不同病理状态下的动态作用机制和调控手段,使其在疫苗设计、肿瘤免疫和自身免疫治疗等领域拥有重要的应用前景。
参考文献:
Ullrich, K. A., Schulze, L. L., Paap, E. M., Müller, T. M., Neurath, M. F., & Zundler, S. (2020). Immunology of IL-12: An update on functional activities and implications for disease.EXCLI journal,19, 1563–1589.
Gerber, A. N., Abdi, K., & Singh, N. J. (2021). The subunits of IL-12, originating from two distinct cells, can functionally synergize to protect against pathogen dissemination in vivo.Cell reports,37(2), 109816.
Liu, J., Cao, S., Kim, S., Chung, E. Y., Homma, Y., Guan, X., Jimenez, V., & Ma, X. (2005). Interleukin-12: an update on its immunological activities, signaling and regulation of gene expression.Current immunology reviews,1(2), 119–137.
Watford WT, Hissong BD, Bream JH, Kanno Y, Muul L, O'Shea JJ. Signaling by IL-12 and IL-23 and the immunoregulatory roles of STAT4.Immunol Rev. 2004;202:139-156. doi:10.1111/j.0105-2896.2004.00211.x
Xiao Z, Casey KA, Jameson SC, Curtsinger JM, Mescher MF. Programming for CD8 T cell memory development requires IL-12 or type I IFN.J Immunol. 2009;182(5):2786-2794. doi:10.4049/jimmunol.0803484
Goriely S, Molle C, Nguyen M, et al. Interferon regulatory factor 3 is involved in Toll-like receptor 4 (TLR4)- and TLR3-induced IL-12p35 gene activation.Blood. 2006;107(3):1078-1084. doi:10.1182/blood-2005-06-2416
Xu, M., Mizoguchi, I., Morishima, N., Chiba, Y., Mizuguchi, J., & Yoshimoto, T. (2010). Regulation of antitumor immune responses by the IL-12 family cytokines, IL-12, IL-23, and IL-27.Clinical & developmental immunology,2010, 832454.
Floss, D. M., Klöcker, T., Schröder, J., Lamertz, L., Mrotzek, S., Strobl, B., Hermanns, H., & Scheller, J. (2016). Defining the functional binding sites of interleukin 12 receptor β1 and interleukin 23 receptor to Janus kinases.Molecular biology of the cell,27(14), 2301–2316.